The New Pandas
Has Creationist Scholarship Improved?
A detailed critique of the new edition of "Of Pandas and People"(1993. Percival Davis and Dean H. Kenyon, Haughton Publishing Company.)
Contents: |
|
Department of Zoology
University of Oklahoma
Norman, Oklahoma, 73019
The New Pandas: A First Look
Frank J. SonleitnerDepartment of Zoology
University of Oklahoma
A second edition of "Of Pandas and People" (Percival Davis and Dean H. Kenyon, Haughton Publishing Co., Dallas, Texas) was published in 1993. The first edition of this book, published in 1989 and intended to be a supplementary biology text emphasizing "intelligent design", was reviewed (Scott 1990, Skoog 1989, Padian 1989, Ruse 1989) and extensively critiqued (Sonleitner 1991). It was found to be sloppily produced with many incidental errors (spelling, grammatical, stylistic, numerical errors in tables, incorrectly drawn and/or labeled figures, errors in citations of references) in addition to gross distortions and errors in the presentation of the biological material. The new edition is little changed from the first. Most of the incidental errors have been corrected, some of the material has been rewritten or replaced but nearly all the distortions and errors of scientific facts remain. Some have even been compounded!
Many of the 130 odd grammatical and stylistic errors compiled by retired high school teacher G. E. Peterson have been corrected. But new ones have been committed! Try out this sentence that begins at the end of page vii: "Within intelligent design also the details as to how gradual or abrupt, and over what span of time differ."
Some biological errors that do not compromise the intelligent design arguments are corrected. Thus amino acids no longer have NH3 groups; turtles are no longer amphibians; the authors grudgingly admit that giraffes might occasionally browse leaves from trees; no longer is it claimed that paleontologists could not distinguish between a placental wolf and a Tasmanian wolf skull—the authors even admit that marsupials might differ from placentals in more ways than just the presence or absence of a pouch. Some of the figures have been redrawn or replaced; those with errors have been corrected. In Figure 4-14 the halloween mask face of the Australopithecus africanus has been fixed, but not the incorrect thorax.
The footnoted references, which were hidden in the Teachers' Guide in the first edition, now appear at the end of each Excursion Chapter. Excursion chapters 1, 2 and 6, which previously didn't have any references, now do, and additional ones have been added to the other chapters. The citations are as sloppy as ever—I have found nearly a dozen mistakes in the new ones.
With a few exceptions to be noted below, the changes in the text are cosmetic. One change that pervades the book is that most of the references to "evolution" and "evolutionists" have been changed to "Darwinism" and "Darwinists". This is apparently to make a distinction between "evolution" which can mean "change in living things over time" and "Darwinism" which refers to the mechanism of mutation and natural selection (Glossary, p. 149, A Note to the Teachers, pp. 155-156.) Changes in living things over time as documented in the fossil record is accepted as a fact (pp. 26, 155-156) and the authors would appear to believe in a kind of progressive creation, although they nowhere specify any such details of their intelligent design hypothesis. I suspect this was done so that creationists of every variety can read their own doctrine into the book.
The Overview chapter has few changes. There has been some shuffling of paragraphs on pp. 18-19 and the last section has been modified to reflect changes in Excursion chapter 6. The partial Dayhoff cytochrome c matrix displayed on p. 37 is free of errors.
In Excursion chapter 1 (the Origin of Life), the section titled: Scientific Case for the Intelligent design of Life, has been rewritten and the authors propose a new law: "Information never arises from physical or chemical causes alone." Because this is really a conclusion based on the assumption of unknowable, untestable, supernatural designers of DNA, it obviously fails the criteria for a scientific law.
In Excursion chapter 2 (Genetics and Macroevolution) the section describing Ambrose' ideas on how evolutionary change might come about has been rewritten, but without changing its grossly inaccurate content. The authors still insist that many features of the giraffe's "adaptational package" are unique to the giraffe. The stone plants and the accompanying figure in the first editions have been omitted.
There are no major changes in Excursion chapter 3 (the Origin of Species). Phillip Johnson's book has been added to the Suggested Reading / Resources.
In Excursion chapter 4 (The Fossil Record), the sections on the origin of reptiles and of plants have been omitted. The discussion of Homo erectus has been greatly shortened and a discussion of the mitochondrial Eve added. There is also a new discussion of the mammal-like reptiles and a more accurate description of Archaeopteryx, but it is claimed that natural selection could not be responsible for the evolution of feathers. The authors mention the new finding that the whale Basilosaurus had tiny hind feet. They do not mention Gish's contention that this specimen is a reptile, but on the basis of Gingerich's speculation that the tiny feet aided in copulation, claim that this is an example of intelligent design. They also cite Stephen Gould as saying that there is not enough time in the fossil record for the evolution of whales from mesonychids.
There are no major changes in Excursion chapter 5 (Homology). The authors omit their previous short discussion of cladistics. Although they continue to couch some of their examples in terms of recapitulation, they now specifically say that recapitulation is false (p. 129).
Excursion chapter 6 (Biochemical Similarities) has undergone the most extensive revision. Sections on the complex mechanism of blood clotting and on the origin of proteins have been added. It is interesting to note that the blood clotting mechanism has been used by Jacob (1977) as an example of "tinkering" by natural selection! To make room for this new material, the section on cytochrome c has been greatly abridged. Any possibility that this data might support evolution (and such a discussion did appear in the first edition) has been eliminated. Now the authors, even though they admit that they are comparing living forms, none of which is ancestral to any other, fly in the face of reason and logic and insist that one should find intermediate degrees of divergence of cytochrome c when comparing carp, bullfrogs, turtles, etc. This misinterpretation of the cytochrome c data was pioneered by Denton (1985) and has been referred to as "Denton's error" by a number of evolutionary authors and reviewers (Landau 1990, Thwaites 1989, Thwaites 1990). According to Davis and Kenyon, the biochemical evidence totally refutes evolution. On this basis alone, I must conclude that the second edition of Pandas is a retrograde development—it is worse than the first edition!
Charles Thaxton's "A Word to the Teacher" has been supplanted by one written by M. D. Hartwig, and S. C. Meyer, in another attempt to make supernatural mechanisms viable scientific explanations.
References:
Denton, M. 1985. Evolution: A Theory in Crisis. Bethesda, MD: Adler and Adler.
Jacob, F. 1977. Evolution and Tinkering. Science 196: 1161-1166 (see p. 1165).
Landau, M. 1990. Protein Sequences and Denton's Error. Creation/Evolution 9(2): 1-7 (Winter).
Padian, K. 1989. Gross Misrepresentation. Bookwatch Reviews 2(11): 2-3.
Ruse, M. 1989. They're Here! Bookwatch Reviews 2(11): 3-4.
Scott, E. C. 1990. Of Pandas and People. NCSE Reports 10(1): 16-17 (Jan.-Feb.)
Skoog, G. 1989. A View From the Past. Bookwatch Reviews 2(11): 1-2.
Sonleitner, F. J. 1991. What's Wrong With Pandas? 106 pp. National Center for Science Education.
Thwaites, W. 1989. Evolution: A Theory in Crisis. NCSE Reports 9(4): 14-17 (July-Aug.)
Thwaites, W. 1990. Molecular Clocks and Creationism. NCSE Reports 10(5): 14-16 (Sept.-Oct.)
The New Pandas: A Closer Look. Is It Improved?
Department of Zoology
University of Oklahoma
December, 1994
A second edition of "Of Pandas and People" (Percival Davis and Dean H. Kenyon, Haughton Publishing Co., Dallas, Texas) was published in 1993. In "A Note to Teachers" included in the back of the book, Mark Hartwig and Stephen Meyer say (p. 154):
"The purpose of this text is to expose your students to the captivating and the controversial in the origins debate ... and encourage them to grapple with ideas in a scientific manner.
Pandas does this in two ways. First, it offers a clear, cogent discussion of the latest data relevant to biological origins ...
Second, Pandas offers a different interpretation of current biological evidence ... a clear alternative, which the authors call "intelligent design."
If Pandas really did this, it would be very nice. But Pandas fails in two ways:
(1) It provides no alternate explanation. "Intelligent design.i.Intelligent design;" is like the emperor's new clothes. Behind the impressive name lurks nothing but a totally mysterious, invisible and inaccessible supernatural agent, a concept whose information content and explanatory usefulness is zero.
(2) In order to persuade the reader to accept this "explanation", Pandas inundates the reader with a flood of disinformation about biology, biologists and evolution.
In "The New Pandas: A First Look", I pointed out that the footnoted references, now appear in the book itself. Also, the Glossary (pp. 149-151) has been expanded; the Acknowledgements section (p. iii) now includes the professional affiliations of the critical reviewers and brief background descriptions of the authors and contributors are given on p. 169.
A pervading change in the book is that most of the references to "evolution" and "evolutionists" have been changed to "Darwinism" and "Darwinists" to make the distinction between "evolution" which can mean "change in living things over time" and "Darwinism" which refers to the mechanism of mutation and natural selection (Glossary, p. 149, A Note to Teachers, pp. 155-156.) Changes in living things over time as documented in the fossil record is accepted as a fact (pp. 26, 155-156.) "Intelligent design" apparently refers to a kind of progressive creation. In regard to this, Pandas mentions the possibility that blueprints of previous organisms might have been used in the creation of later ones (p. 42).
Is the new edition improved? Only a few minor errors and shortcomings of the 1st edition have been corrected. The great majority of the errors, extensively critiqued in "What's Wrong with Pandas?" still remain. The new material is just as unreliable and will be dealt with below. Pandas has changed its discussion of the cytochrome c data (Overview and Excursion chapters 6) to reinforce its incorrect view that such biochemical data disproves evolution. In this respect, the new edition is worse than the first.
Pandas' Introduction
This has been rewritten, involving only cosmetic changes.
Of Pandas and People: An Overview
Overview Section 1: The Origin Of Life
On p.4 Pandas says that there is geological evidence that there was an oxygen atmosphere earlier than the development of life. This is not correct. There is evidence for small amounts of oxygen in the atmosphere of later Precambrian times as a result of the photosynthetic activities of some of the prokaryote organisms.
Figure 1 has been corrected. The amino acid structures now have NH2 groups instead of NH3 groups.
Overview Section 2: Genetics And Macroevolution
In column 1 of p. 12, Pandas now say that some mutations may be neutral. In column 2, they admit that giraffes feed in tree tops. In column 2 of p. 13, Pandas cites brachiopods (see p. 72 for reference) as another example of interdependence of structures. There is nothing particularly special about brachiopods in this regard.
Overview Section 3: The Origin Of Species
On pp. 18-19, some of the paragraphs have been shuffled around between the sections on Genetic Drift and The Founder Effect.
Overview Section 4: The Fossil Record
The "avian complex" (p. 23, column 2, top) will be discussed in connection with Excursion Chapter 4.
Facts Versus Interpretation
Here Pandas states: "The existence of fossils with enormous variety is a fact, and so are the changes in the distribution of those fossils over time;..." If we accept that all life comes from life, as Pandas insists then the younger fossils must have descended from the older ones, and we have a documented evolutionary history! Only if we assume spontaneous generation by intelligent designer intervention, can we reconcile the intelligent design hypothesis with these facts. And this produces a kind of "progressive creationism" as described by Ambrose (1982), somewhat akin to theistic evolution!
Overview Section 5: Homology
There is a new and improved (really) Figure 6 on p. 28.
Contradictory Similarities
Here Pandas admit that the American wolf and the Tasmanian wolf differ in more characters than just the presence or absence of a pouch and the statement that a paleontologist could not distinguish between fossils of the two has been deleted.
Overview Section 6: Biochemical Similarities
The introductory paragraph has been rewritten. There is a new figure on p. 36. The Dayhoff matrix in Table 1 (p. 37) has been redone and the 6 paired errors in the 1st edition corrected.
A New Pattern
In the 1st edition, Pandas spoke of a "ladder" of evolution (column 1, p. 37.) Now Pandas notes that evolution follows a branching tree pattern (column 2, p. 37). Yet Pandas still insists that the living forms included Table 1 should display intermediate cytochrome c patterns if Darwinism is correct! There is no logical reason for this assertion! Even in terms of anatomy, the bullfrog, for example, is a highly specialized amphibian and is not intermediate between the fishes and reptiles. For the true intermediate forms one must look to the fossil amphibians in the Paleozoic.
The Molecular Clock
This was originally a part of the preceding section. The subject has been eliminated from Excursion Chapter 6.
The Task Of Taxonomy
This section has been extensively rewritten. Pandas' claim that the fossil record also shows distinct clusters (p. 39 bottom of column 2) is not true.
"The amphibians grade so insensibly into the reptiles that the assignment of certain fossils becomes rather arbitrary...The same is true of the border between mammal-like reptiles and the mammals." (Mayr, 1963, p. 596.)
References
Ambrose, E. J. 1982. The Nature and Origin of the Biological World. Halsted Press. New York.
Mayr, E. 1963. Animal Species and Evolution. The Belknap Press of Harvard University Press. Cambridge.
Excursion Chapter 1: The Origin Of Life
There are only a few minor changes. The amino acid structures in Figure 1-5 (p. 51) have been corrected to have NH2 groups instead of NH3 groups.
Problems With The Assumptions Of The Oparin Hypothesis
Pandas' reference 6 (Clemmey and Badham, 1982) summarizes the evidence for oxygen in the Precambrian atmnosphere. Before the advent of life, free oxygen might have been generated through photodissociation of water by ultra-violet rays. They also mention that several of the ‘origin of life' scenarios: prebiotic evolution in space, possibly in comets, and among pores in the dust grains of the early Earth regolith, do not require a reducing atmosphere.
Scientific Case For The Intelligent Design Of Life
Here Pandas claims that "Information never arises from physical or chemical causes alone" is an empirically based generalization or law of nature. This is supposed to be based on "our universal experience that such sequences are the result of intelligent causes" (p. 58). But this is not the case. It is not our empirical experience that supernatural designers have created DNA! Intelligent design proponents can only postulate that DNA sequences were produced by intelligent design. In fact, in the section "A Note to Teachers" (see p. 160), this generalization is said to be a prediction of the intelligent design hypothesis! Pandas is indulging in circular reasoning. The generalization upon which they base their hypothesis is itself based on that hypothesis.
An Analogy
Towards the end of this section (bottom of p. 57), Pandas advises, "Well-designed experiments on the origin of life should continue." One might ask, "Why?" If life originated as the result of a supernatural, intelligent designer, no experiments will ever yield any understanding of the phenomenon. Maybe Pandas knows something but they're not telling us. What do Pandas consider to be a well-designed experiment? And what possibly could the experimenter be looking for?
References
Clemmy, H. and N. Badham. 1982. Oxygen in the Precambrian atmosphere: An evaluation of the geological evidence. Geology 10(3): 141-146.
Excursion Chapter 2: Genetics And Evolution
Nothing of importance has been changed in this chapter. Table 2-1 now has a legend. The term "messages" is substituted for "Specifications" in Figure 2-3. The discussion of the "stone plants" and the accompanying Figure 2-6 have been omitted. The book by Yockey listed under Suggested Reading/Resources now has a 1992 publication date but the title is wrong! The correct title is "Information Theory and Molecular Biology."
Certain parts of the chapter have been more or less rewritten but without changing any arguments or adding anything new.
Change And The Origin Of New Structures
This section, which incorporates Ambrose' ideas (Ambrose, 1982) on the nature of evolutionary change has been extensively rewritten but without correcting any of the grossly inaccurate content. Ambrose claims that the application of information theory disproves evolution (p. 69, top of 2nd column). Yockey (1992) disagrees with this and essentially corroborates what I previously said about evolutionary change.
Natural Selection And The Adaptational Package
In two place in column 1 of p. 71, Pandas admits that giraffes might use their long necks to browse in the trees!
References
Ambrose, E. J. 1982. The Nature and Origin of the Biological World. Halsted Press. New York.
Yockey, H. P. 1992. Information Theory and Molecular Biology. Cambridge University Press, Cambridge.
Excursion Chapter 3: The Origin Of Species
There are only a few minor changes in this chapter that require comment.
The unnumbered figure at the top of p. 82 has two labels "Original Gene Pool" and "New Gene Pool" added to it. There is a new version of Figure 3-4 (on p. 84). Phillip Johnson's book, Darwin on Trial, has been added to the Suggested Reading/Resources.
Near the bottom of the first column on p. 78 Pandas gives an example of a phylum with only one species, but the names are reversed! It should be the phylum Caryoblastea containing the one species Pelomyxa palustris. One might rightly guess that these are highly unusual organisms! They are giant single cells visible to the naked eye and the most primitive of eukaryotes. They have membrane-bound nuclei but no endoplasmic reticulum, Golgi bodies, mitochondria, chromosomes or centrioles. They seem to divide as bacteria do; there being no sign of a mitotic process (Margulis and Schwartz, 1982).
In column 2 of p. 78, Pandas says that "the challenge is discovering the extent of change that takes place..." I think the real challenge is discovering how species were designed in the first place!
The Failure Of Natural Selection
Pandas still hasn't got the quotes straight! Mayr's statement (Mayr, 1963, p. 12) is attributed to Simpson (Simpson, 1964, p. 81) and vice versa.
In the 2nd column of p. 88, Pandas has a new sentence:
"Moreover, a growing number of scientists accept natural selection as a reasonable explanation for the modification of traits but not for the origins of new structures."
and gives 6 citations (reference 5) to back it up. Although this statement paraphrases Rieppel (1990, p. 303), I think the sentence is misleading. Whether we are dealing with a modified trait or the appearance of an incipient structure, we are dealing with variation which originates from mutation and/or genetic recombination. Natural selection only preserves modifications and new structures, guiding the course of evolution by controlling the accumulation of such variations and combinations of them through elimination or preservation. Reippel cites Sober (1984, p. 197) who suggests that the dorsal plates of Stegosaurus originated fortuitously. Once they occurred they may have performed a number of adaptive functions and preserved by natural selection. On the other hand, several of the authors cited have difference ideas about the source of variation. Margulis (Mann, 1991, p. 379) thinks it results from the acquisition of symbionts; McDonald (1983, p. 97) postulates stress-induced variation; Ambrose (1982, p. 131, but esp. 143) postulates a designer tinkering with the DNA during speciation by punctuated equilibrium.
References
Ambrose, E. J. 1982. The Nature and Origin of the Biological World. John Wiley and Sons. New York.
Mann, C. 1991. Lynn Margulis: Science's Unruly Earth Mother. Science 252: 378-381. (19 April).
Margulis, L. and K. V. Schwartz. 1982. Five Kingdoms: An Illustrated Guide to the Phyla of Life on Earth. W. H. Freeman and Co., San Francisco.
Mayr, E. 1963. Animal Species and Evolution. The Belknap Press of Harvard University Press.
McDonald, J. F. 1983. The Molecular Basis of Adaptation: A Critical Review of Relevant Ideas and Observations. Annual Review of Ecology and Systematics 14: 77-102.
Reippel, O. 1990. Structuralism, Functionalism, and the four Aristotelean Causes. Journal of the History of Biology 23(2): 291-320.
Simpson, G. G. 1964. This View of Life. The World of an Evolutionist. Harcourt, Brace and World.
Sober, E. 1984. The Nature of Selection: Evolutionary Theory in Philosophical Focus. University of Chicago Press. Chicago and London.
Excursion Chapter 4: The Fossil Record
Modifications to this chapter include the deletion of the sections on the origin of reptiles and plants (Figure 4-12 from the 1st edition has been deleted) and a rewriting of the sections on the origin of mammals and Archaeopteryx. The material on Homo erectus which was mostly wrong has been greatly reduced and a section on the mitochondrial Eve added.
Major Features Of The Fossil Record
Feature No. 1 (that fossils encapsulate information about past, extinct forms) has been omitted from this section and the following one.
Early Expectations Of The Fossil Record
In this section, Figure 4-2 has been revised. Only animal phyla are shown (Pandas still doesn't identify them) thereby omitting a number of phyla (mostly plants) whose origins either predate the beginning of the Cambrian or occur much later. This bias accentuates the "Cambrian explosion." The figure has been marginally improved by indicating the known fossil record (solid line) and presumed fossil record (dotted line) of each phylum, showing quite clearly that the origin of most of the animal phyla is only presumed to have occurred at the beginning of the Cambrian.
Gaps And Grouping In The Fossil Record
Mammals
This section has been rewritten and is fairly accurate. Pandas even admits that, with regard to the mammal-like reptiles, "Indeed, it does appear that they provide Darwinists with a superior example of a transitional series." (Pandas, p. 100)
Actually they provide all evolutionists, including non-Darwinists such as Grasse (Grasse, 1977) and even "progressive creationists" such as Richard Owen (Desmond, 1982), with this superior example! Pandas' speculation that the information for mammalian structures "may have existed, unexpressed, in the genome of the earliest Therapsids before they diversified" is pure speculation and would involve totally unknown genetic mechanisms. It is not clear why Pandas bothers to make this speculation, unless it is to convince one that the evolution of Therapsids is simply "horizontal variation."
"The absence of unambiguous transitional fossils" is no longer illustrated by whales. Gould (1994) summarized all but the latest remarkable transitional whale fossils found in the past few years, including Pakicetus, the amphibious mesonychid-like form, Ambulocetus and Indocetus which are four-legged whales without modified tails, and the new Basilosaurus with the tiny regressed hind legs. Gould obviously believes that whales evolved from mesonychids! How is this fact reconciled with Pandas' claim that Gould has calculated that there was not enough time to evolve a Basilosaurus from the Mesonyx by punctuated equilibrium? The claim is false. Steven Stanley argued that there wasn't time to evolve the whales from mesonychids by phyletic gradualism but there wouldn't be any problem if most of the evolution occurred by punctuated equilibrium at speciation events (Stanley, 1981, pp 93-95). This argument is discussed both by Augros and Stanciu (1987, p. 175) and by Johnson (1991, p. 51) which is probably where Pandas encountered it but its presentation here is hopelessly garbled.
The latest whale find is the 46 million year old Rodhocetus, whose legs were shorter than those of Ambulocetus, its sacral vertebrae not fused and not articulating with the pelvis and whose massive tail vertebrae indicate that it had powerful tail flukes (Gingerich et al, 1994; Zimmer, 1995) thus filling a gap between Ambulocetus and Basilosaurus. We now have a complete set of fossils documenting the major steps in whale evolution! Of course it is not fair to criticize Pandas for not mentioning discoveries that were announced after Pandas had gone to press, but this situation illustrates that creationists, whose ideas rely on gaps in the fossil record, are fighting a rear-guard action which they are losing.
Amphibians
Paleontology is continually finding new fossils that shed light on the fish-amphibian transition. Thus Pandas account of the origin of amphibians is years out of date. Recent publications discuss fossils that are closing the gap between fishes and amphibians. Certain crossopterygian (sarcopterygian) fishes of the family Panderichthyidae are so similar to the earliest amphibians, Ichthyostega and Acanthostega, that, in some instances, it is difficult to tell if a fossil is a fish or an amphibian (Ahlberg, 1991; Ahlberg and Milner 1994.) Acanthostega had fishlike internal gills and ventilation. The limbs had supernumerary digits and were flipper-like; the pelvis was similar to that of a fish (Gee, 1991). On the other hand, the paired fins of the crossopterygian fish Sterropterygion brandei show tetrapod-like features including details of the shoulder girdle; a primitive elbow joint in the pectoral fins and a primitive tarsal joint in the pelvic fins (Rackoff, 1980).
The evolution of amphibians from fish occurred in a chunk of space-time including all the continents of the earth over a period of 28 million years. We have fossils from only a dozen or so specific points in space and time during that entire period so it is not surprising that there are gaps in our knowledge of that process.
Archaeopteryx
Based on its feathers, Pandas claims "that Archaeopteryx was an accomplished flyer, because they provided an airfoil superbly adapted for flight." The superb airfoil is a reference to the fact that the feathers have a vane asymmetry similar to that of extant birds which gives them a wing-like cross-section. A recent study (Speakman and Thomson, 1994) has shown that "the extent of asymmetry in the feathers of Archaeopteryx is only slight, significantly lower than in modern flying birds and comparable to that of extant nonflying birds." Other studies have shown that Archaeopteryx' wrist lacks all the key features for flight found in the modern bird (Vazquez, 1992) and that the flight muscles were too small to provide powerful sustained flight (Speakman, 1993). Thus the evidence shows that Archaeopteryx did not have fully modern feathers and was most likely a very weak flyer. Lacking the full "avian complex" Pandas claims it couldn't fly at all; hence natural selection couldn't have evolved the feather. But if it wasn't "intended" to fly, why did the "intelligent" designer give it the "superb airfoil"?
Evolutionists explain the intermediate nature of Archaeopteryx by saying that it is a transitional form in the evolution of birds. Design proponents have no explanation for its intermediate form except that some unknown and unknowable supernatural designer desired to make it so! Recently, somewhat younger fossil birds have been found from the Lower Cretaceous period that are intermediate between Archaeopteryx and modern birds (Kurochkin, 1985; Sereno and Chenggang, 1992; Sanz, Bonaparte and Lacasa, 1988). The oldest of these, the sparrow-sized Sinornis from China is about 15 million years younger than Archaeopteryx. It looks very much like a small Archaeopteryx, but it has a shorter body, a much shorter tail, a stronger pectoral girdle and forelimbs for much more powerful flight; the wing has an advance wing-folding mechanism and the hind feet are modified for perching. About 5 million years younger than Sinornis is the Spanish Las Hoyas bird which is slightly more modern in form. And the Mongolian Ambiortus, about 5 million years younger than the La Hoyas bird, is even more modern! This is a wonderful series of intermediate forms representing an evolutionary series in the evolution of the "avian complex." Do the design proponents dare to suggest that they are trial efforts of a much less than omnipotent and omniscient designer who has to produce new designs in stages?
In Figure 4-10, the drawing of an unspecified Triassic thecodont has been replaced with one of an unspecified Theropod dinosaur.
The Origin Of Man
Corrections have been made to Figures 4-12 thru 4-14 (formerly Figures 4-13 thru 4-15). No longer does Figure 4-12 show Pliopithecus and Oreopithecus persisting to the present time; numbers for the cranial capacities of A. boisei and A. robustus have been added to Figure 4-13 and the halloween mask skull of Australopithecus africanus in Figure 4-14 has been replaced. In the text, almost all the discussion of Homo erectus, which was fraught with errors, has been eliminated.
Added to this section is a discussion of the mitochondrial Eve hypothesis. Unfortunately it gives a muddled explanation for the use of mitochondrial DNA. Consider this sentence, referring to mutations in mitochondrial DNA: "The reason they do is that such mutations are not corrected as they are in nuclear DNA." What is that supposed to mean?
Actually, mitochondrial DNA is useful for "tracking the relationships among peoples" because it is transmitted to offspring maternally, i.e. only through their mother. As generations go by, however, the mitochondrial DNA of any females who produce only sons is lost, and eventually, the mitochondrial DNA of the species will be reduced to one lineage stemming from one ancestral female. This is the mitochondrial Eve. It definitely does not mean that all races of humanity owe their origin to a single female ancestor. They owe their origin to an entire population of females and males. It is just that the mitochondrial DNA of only one of those females has survived to the present. To make sure that all possible errors have been committed, Pandas reports the citation (reference 23) incorrectly. It should be A. C. Wilson et al.
The quote from Thomson (reference 24: Thomson, 1982) stops just short of where Thomson presents a solution to breaking the "circularity in the approach"—which is to use cladistic analysis! Clever (but dishonest) Pandas! Is this why they eliminated their discussions of cladistics from the new edition?
Design adherents, in claiming that Homo erectus and other hominids are just apes and that culture appeared abruptly, are only displaying their ignorance of the relevant facts. The anatomical evidence is clear that the hominids are, to various degrees, pre-men; the youngest specimens of H. erectus grade into modern man through presapiens (archaic sapiens) forms. And culture first appeared in a very simple form, called Oldowan, dating from 500,000+ years ago and associated with the early Homo species, and developed through a sequence of stages of ever increasing complexity: Acheulian (75,000—150,000 years ago, associated with H. erectus), Mousterian (45,000 years ago associated with Neandertal man), and finally Aurignacian (40,000 years ago), Perigordian (30,000 years ago), Solutrian (20,000 years ago), Magdalenian (15,000 years ago) and Azilian (10,500 years ago) associated with Cro-Magnon (modern) man (Howell, et al, 1965).
References:
Ahlberg, P. E. 1991. Tetrapod or near-tetrapod fossils from the Upper Devonian of Scotland. Nature 354: 298-301 (28 November).
Ahlberg, P. E. and A. R. Milner. 1994. The origin and early diversification of tetrapods. Nature 368: 507-514. (7 April).
Augros, R. and G. Stanciu. 1987. The New Biology: Discovering the Wisdom in Nature. New Science Library. Shambhala. Boston and London.
Desmond, A. 1982. Archetypes and Ancestors: Palaeontology in Victorian London 1850-1875. University of Chicago Press. Chicago.
Gee, H. 1991. The brave vertebrate venture. Nature 354: 268-269. (28 November).
Gingerich, P. D., S. M. Raza, M. Arif, M. Anwar and X. Zhou. 1994. New Whale from the Eocene of Pakistan and the origin of cetacean swimming. Nature 368: 844-847 (28 April). See also: Novacek, M. J. 1994. Whales leave the beach. Nature 368: 807 (28 April).
Gould, S. J. 1994. Hooking Leviathan by Its Past. Natural history 103(5): 8-15. (May).
Grasse, P.-P. 1977. Evolution of Living Organisms: Evidence for a New Theory of Transformation. Academic Press, New York.
Howell,F. C. and The Editors of LIFE. 1965. Early Man. Time Inc., New York.
Johnson, P. E. 1991. Darwin on Trial. Regency Gateway, Washington, D. C.
Kurochkin, E. N. 1985. A True Carinate Bird from Lower Cretaceous Deposits in Mongolia and Other Evidence of Early Cretaceous Birds in Asia. Cretaceous Research 6(2): 271-278.
Rackoff, J. S. 1980. The Origin of the Tetrapod Limb and the Ancestry of Tetrapods. In: Panchen, A. L. (Editor). 1980. The Terrestrial Environment and the Origin of Land Vertebrates. Academic Press. London, New York. pp. 255-292.
Sanz, J. L., J. F. Bonaparte and A. Lacasa. 1988. Unusual Early Cretaceous birds from Spain. Nature 331: 433-435. (4 February). See also: Cracraft, J. Early evolution of birds. Ibid. 389-390.
Sereno, P. C. and R. Chenggang. 1992. Early Evolution of Avian Flight and Perching: New Evidence from the Lower Cretaceous of China. Science 255: 845-848. (14 February). See also: Barinaga, M. 1992. Evolutionists Wing It With a New Fossil Bird. Ibid. 796.
Speakman, J. R. 1993. Flight capabilities in Archaeopteryx. Evolution 47(1): 336-340.
Speakman, J. R. and S. C. Thomson. 1994. Flight capabilities of Archaeopteryx. Nature 370: 514. (18 August).
Stanley, S. M. 1981. The New Evolutionary Timetable: Fossils, Genes, and the Origin of Species. Basic Books, Inc., New York.
Thomson, K. S. 1982. Marginalia: The meanings of evolution. American Scientist 70(5): 529-531. (September-October).
Vazquez, R. J. 1992. Functional Osteology of the Avian Wrist and the Evolution of Flapping Flight. Journal of Morphology 211: 259-268.
Zimmer, C. 1995. Back to the Sea. Discover 16(1): 82-84. (January).
Excursion Chapter 5: Homology
There are only a few minor changes in this chapter. All the old errors are still intact.
Pandas appears to have some difficulty with the concepts of parallel and convergent evolution. In the first edition, on p. 117 they claimed that the American wolf and the Tasmanian wolf are examples of parallel evolution, the vertebrate eye and the squid eye are an example of convergent evolution and on p. 120, the common features of the two pandas are the result of parallel evolution. In the second edition, the two terms are interchanged, the wolves and the pandas are said to represent convergent evolution and the vertebrate and squid eye parallel evolution! Whether similarities are due to convergence or parallelism depends on the closeness of relationship of the forms involved. Thus the vertebrate and squid eyes are definitely a case of convergence, the wolf-like forms are convergent and the pandas may or may not be considered convergent (the Ursidae (bears) and Procyonidae (racoons) are considered close relatives.)
In the second columns of pp. 127 and 133, Pandas omit their previous discussion of cladists.
Some people have claimed that the panda's thumb is incorrectly labeled in Figure 5-6. Labeled diagrams of the panda's hand (Gould, 1978; Tabin, 1992) indicate that Figure 5-6 is correct! (How did Pandas manage that?) In addition to asking why the intelligent(!) designer gave the panda an ersatz thumb instead of a true opposable thumb, such as the primates possess, why is it that the panda's foot also has an enlarged tibial sesamoid, but not enlarged enough to be a "panda's big toe"? (Gould, 1978, p. 30; Roth, 1988, p. 21).
Homologies And The Fossil Record
Here Pandas mention the mitochondrial DNA of some 20 million year old trees as an unusual type of fossil. In the reference cited, Gould (Gould, 1992) discusses the DNA extracted from leaves of Magnolia and Taxodium (the bald cypress) recovered from Miocene lake beds in Idaho. Gould's paper is subtitled "A colorful 20-million-year-old leaf is an elegant test of the theory of evolution." The changes in gene sequences between these fossils and their modern counterparts is a remarkable confirmation of evolution, involving the accumulation of neutral mutations, and by implication, a confirmation of molecular clocks.
What Is Homology, Really?
Virtually all the homologies of vertebrate comparative anatomy were identified by early 19th century pre-evolutionary anatomists. Most of these structures not only share common connections and relations with other structures but also a common embryological origins. But there are exceptions which were discussed and summarized by De Beer (1958, 1971). De Beer's 1971 monograph served as the basis for chapter 7 of Denton (1985) and is mentioned by Johnson (1991) in his Research Notes to Chapter 5 (p. 172). These authors have misread de Beer to mean that no homologous structures have similar embryological origins or are even determined by the same genes! Denton concludes that homology is a useless concept although de Beer did not! (de Beer, 1958, p. 152).
These problems have been further considered by Roth (1984, 1988) and in a new book on homology (Hall, 1994). Complexities arise because there is not a simple congruence between the genes, developmental mechanisms and morphological features. Not only can a morphological feature evolve, but also the developmental mechanism producing it. Complexes of pleiotropic genes interact by way of a hierarchy of interrelated developmental mechanisms to produce the morphological phenotypic character. The recent discovery of homeotic (homeobox) genes governing the anterior-posterior differentiation of a wide variety of organisms has revealed an extensive homology within their genetic regulatory systems, and has helped to explain in part some of the difficulties mentioned by de Beer. For more on homeotic genes, see the new supplement: Homology in Developmental Genetics: the homeotic genes.
The Coccyx: An Easy Case For Evolution
At the end of this section in the first edition, Pandas admitted that the coccyx "seems to be homologous to a tail." If this is accepted then, as a tail, the coccyx is definitely vestigial.
Recapitulation: An Evolutionary Application Of Homology
Near the top of the second column on p. 129, Pandas has added some sentences indicating that the biogenetic law has been rejected by science. In the paper cited (Thomson, 1988), Thomson makes clear that it is the Meckel-Serres and Haeckelian type of recapitulation that has been rejected. On the other hand he says, "Von Baer and his predecessors had hit upon a fundamental fact of nature" and later, "Although von Baer was an avowed anti-Darwinist, what has become known as "von Baerian recapitulation" is wonderful evidence for the fact of evolution.." Interestingly enough, nowhere in this paper do I find the statement attributed to Thompson by Pandas that "The biogenetic law as a proof of evolution is valueless."
Raff and Kaufman (1983) do say that the biogenetic law is "a mirage" on p. 19, but later in the book (chapter 5, especially pp. 154 fol. and chapter 6) they give a modern explanation of von Baer's laws, which is the same as that given in the "What's Wrong With Pandas" supplement, "Embryos and Evolution." It would seem that Pandas don't read the references they cite, because Raff and Kaufman (1983) is a very good summary of the interrelationships of embryology and evolution, containing much important evidence for evolution, including discussions of punctuated equilibrium, molecular clocks, how homeotic genes govern body plans and the two chapters mentioned dealing with the true nature of recapitulation phenomena.
The Kidneys: An Easy Case For Recapitulation
At the end of this section, Pandas states, "...scientists have recently recognized that embryonic structures once thought functionless actually play important roles in the formation of more advanced structures to follow." The example given in the citation (Raff and Kaufman, 1983, p. 18) dates back to 1886—more than 108 years ago—which is hardly recent!
The Intelligent Design Interpretation Of Homology
Although Stephen Gould may have said that homology supports common design as well as it does common ancestry, he does not say it in the paper cited (Gould, 1987).
References:
de Beer, G. R. 1958. Embryos and Ancestors. 3rd Edition. Clarendon Press, Oxford.
de Beer, G. R. 1971. Homology: an Unsolved Problem. Oxford Biological Readers, No. 11. Oxford University Press. London.
Denton, M. 1985. Evolution: A Theory in Crisis. Adler and Adler. Maryland.
Gould, S. J. 1978. The Panda's Peculiar Thumb. Natural History LXXXVII(9): 20-30 (November).
Gould, S. J. 1987. The Panda's Thumb of Technology. Natural History 96(1): 16-23. (January).
Gould, S. J. 1992. Magnolias from Moscow. Natural History 101(9): 10-18 (September).
Hall, B. K. (Editor) 1994. Homology: The hierarchical basis of comparative biology. Academic Press. New York.
Johnson, P. E. 1991. Darwin on Trial. Regency Gateway. Washington, D. C.
Raff, R. A. and T. C. Kaufman. 1983. Embryos, Genes, and Evolution: The Developmental-Genetic Basis of Evolutionary Change. Macmillan Pub. Co., Inc. New York.
Roth, V. L. 1984. On homology. Biological Journal of the Linnean Society 22: 13-29.
Roth, V. L. 1988. The Biological Basis of Homology. In: Humphries, C. J. (Editor). Ontogeny and Systematics. Columbia University Press. New York. pp. 1-26.
Tabin, C. J. 1992. Why we have (only) five fingers per hand: Hox genes and the evolution of paired limbs. Development 116(2): 289-296 (October).
Thomson, K. S. 1988. Marginalia: Ontogeny and phylogeny recapitulated. American Scientist 76(3): 273-275 (May-June).
Excursion Chapter 6: Biochemical Similarities
Because this chapter has been extensively rewritten, it is appropriate that a new outline be given:
Outline of the Pandas Chapter
All but 4 of the original 16 figures have been deleted. Of the those remaining:
- old Figure 6-1 becomes new Figure 6-1,
- old Figure 6-5 becomes new Figure 6-2, and
- old Figure 6-8 becomes new Figure 6.3.
The new Figure 6-1 has "Organelles" corrected labeled as "Rough endoplasmic reticulum." Recall that the old Figure 6.6 was a Dayhoff matrix (similar to Figure 6 (p. 37 in the Overview section) except that it included 25 species and contained 13 sets of paired errors! It is now gone. A new Figure 6-4 is similar to the old Figures 6-12 and 6-13. New Figures 6.5 and 6.6 illustrate the new section on blood clotting. Old Table 6.1 has been deleted.
The Structures Of Molecules
This section has been partly rewritten. The paragraph on chirality (asymmetry or handedness of molecules) has been deleted.
Common Pathways, Common Code
The paragraphs explaining the genetic code have been deleted.
Similarities In Molecular Sequences
This section has been greatly abbreviated. Subsections on Family Trees, The Search for Intermediate Sequences, The Data Fit a Pattern, the Molecular Clock Hypothesis and Multiple Molecular Clocks have been deleted. All discussion of how evolution explains these data has been stricken, although a section on molecular clocks still exists in the Overview Section (p. 39) as a "vestige" from the 1st edition.
In contrast to Pandas who insist that evolution cannot possibly explain the cytochrome c data, Dayhoff, Park and McLaghlin (1972) state, "From protein sequence data alone, it is possible to derive a phylogenetic or evolutionary tree which shows in detail the nature of the ancestral relationships of present-day species." They present such a tree in their Figure 2-1 and say, " This tree...has the same topology (order of branching without regard for changes in the length of branches or in the angles between them) as trees derived from morphological or other biological considerations." In making the tree, they "use a method that involves reconstructing plausible ancestral sequences from our knowledge of present-day sequences." From their published sequences of cytochrome c from living forms and that of the postulated common ancestors (foldout on p. D-367), I have produced Figure 6.1 which is to be compared with Pandas' Figure 9 (p. 38) or 6-4 (p. 140). The common ancestral sequences are represented by the nodes numbered 2—6. The numbers beside the branches of the tree are the number of amino acid changes occurring along those branches. Examination of this tree (and of the original sequences in Dayhoff, 1972) illustrate that nodes 2 and 3 are intermediate between the horse (or rabbit) and node 6 (the common ancestor of the vertebrates.) The same holds true for nodes 3 and 4. Yet the total number of changes between node 6 and the horse is 13, comparable to the 15 between node 6 and the carp.
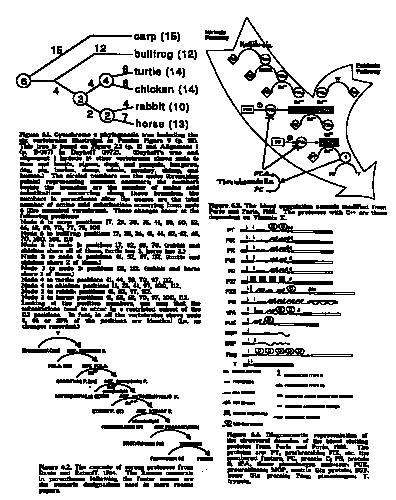 |
All the changes in cytochrome c are neutral, so they continue to accumulate throughout time along each branch, resulting in the fact that none of the living forms at the ends of the branches are intermediate between any of the other living forms. The fact that most of the animals located above a node share the substitutions that occur below that node is strong evidence that those animals are descended from a common ancestor. In general, the closer two organisms are in the classification, the more substitutions they have in common, exactly what one would expect on the basis of evolution. On the other hand, because these substitutions are neutral, there is no possible reason why an intelligent designer should want to make such changes. Under the intelligent design hypothesis, all the cytochrome c proteins should be identical!
A Biochemical System At Work
A Characteristic Of Intelligent Design
Creeping Toward Clotting
Pandas reveals the rationale of its design argument in these sections. Biochemical systems are composed of many interactive parts which have to operate simultaneously. Leaving out any component leaves the system inoperative. Thus it is impossible that they could have evolved in a step-wise fashion; they must have been created in their fully complex form by intelligent designer(s).
But there is something wrong with this argument. Let us apply it, as Pandas does, to an analogous system—the automobile. Modern cars are very complex pieces of machinery, with complex electrical systems, pollution controls, and computer control of many aspects of the engine's operation. Problems with any of these systems may render the car inoperative. They must all operate simultaneously. Therefore it follows that no simpler designs of cars are possible. The car that Henry Ford invented at the turn of the century must have had all these systems in their present degree of complexity. Of course, that is not true.
When a new component is added to a system, the old components which formerly worked by themselves in the old version of the system, have their interactions with the system modified and possible made dependent on the new component. The same could very well have happened in the evolution of the clotting system. Pandas "logical" argument for a designer is faulty.
The Mechanism Of Blood Clotting
Localization Of The Blood Clot, Strengthening, And Dissolution
Pandas' description of the blood clotting mechanism is so awkward and the accompanying figure (Figure 6-6) so poorly designed that the mechanism appears much more complicated than it really is. My Figure 6.2 emphasizes the sequential pattern of serum proteases that form a cascade and amplifying mechanism. Figure 6.3 contains a more modern and detailed presentation. Each of the active enzymes has a short mean life span, but as the enzymes in each stage produce a larger number in the following stage, one molecule of Hageman factor may result in thousands of molecules of fibrin being produced in a very short time. The system acts as an amplifier (McFarlane, 1964; Davie and Ratnoff, 1964). The figures also shows the factor numbers that have been used in place of names in the more recent literature. Thus Hageman factor is Factor XII, etc.
This cascade could easily have evolved from the bottom up, the insertion of each additional step enhancing the amplification and fine-tuning of regulation (Neurath, 1984). The greater blood pressures and activity of higher animals may have been the selective factor in increasing the haemostatic efficiency of this mechanism (McFarlane, 1964). Factors XII (Hageman factor), prekallikrein and HMWK (high molecular weight kininogen) at the upstream end of the intrinsic pathway can be defective and the clotting mechanism still works (Furie and Furie, 1988). Thus Pandas' statement "all of the proteins had to be present simultaneously for the blood clotting system to function" (top, column 1, p. 146) is wrong.
Figure 6.4 shows diagrammatically the various homologous domains these proteins share, indicating how they could have evolved via gene duplication, modification and exon shuffling (Furie and Furie, 1988). Based on these structures, Patthy (1985) has reconstructed their evolutionary history. The catalytic domain of the serum proteases is almost identical to that of the digestive proteases, such as trypsin. Their inhibitors also show striking homology to the inhibitors of the digestive proteases. These homologies suggest common ancestry for all these proteases (Neurath, 1984). This development of the clotting system from other preexisting proteins is a example of "tinkering" by natural selection, rather than the activity of a conscious, intelligent designer (Jacob, 1977).
Nature's Search For A Protein
Pandas cite Yockey (1977, not 1978!) as having published calculations on the origin by chance of a 100 amino acid protein. Actually, he considered cytochrome c with 101 amino acids and arrived at a probability of 2.1 x 10-65. This was done in the following manner: Given 20 different amino acids, and making certain allowances, there are 1.8067 x 10126 possible sequences of 101 amino acids. Based on the chemical similarity of various amino acids and hence the possibility of neutral substitutions, there are 3.8 x 1061 different cytochrome c sequences, or 3.8 X 1061/1.8067 x 10126 = 2.1 x 10-65 of the total (Yockey, 1977, pp. 386-387).
In a similar manner, Reidhaar-Olson and Sauer (1990, p. 315—Pandas' reference 5), on empirical grounds, estimated that there were 1057 functional sequences of a particular 92-residue domain, or 1 x 1057/4.9517 x 10119 = 2 x 10-63 of the total, a proportion similar to Yockey's.
On the other hand, Lau and Dill (1989, p. 3993—Pandas' reference 4), assuming there are only two kinds of amino acid residues (hydrophobic and polar), calculated the folding patterns for all possible sequences of 1024 residues. Six (5.86 x 10-03) had only 1 native state (folding pattern or conformation), which might confer a biological advantage compared to others that exhibited a number of conformations. They suggest that if there were 20 types of residues, there might be a larger population of sequences with only one or a few conformations.
Underlying these calculations is the assumption that, for example, the first cytochrome c had 101 positions and worked in same fashion as the cytochrome c of today. Maybe it was much simpler. An investigation of the family of calcium-binding proteins suggests that a modern protein with 160 amino acids originally only had 40 (Awbrey and Thwaites, 1981, p. 16). Fully a third of all proteins and enzymes are built around an inorganic metal atom-containing prosthetic group, such as the porphyrin ring found in cytochromes, hemoglobins, and chlorophyll. The prosthetic group can perform the function of the protein by itself. The protein simply fine tunes the action of the prosthetic group to make it more specific and efficient (Anon. 1980). Such proteins could have originated from very short amino acid chains that grew larger with time. Finally, there is evidence that many large proteins originated as many repetitions of short amino acid oligomeres (Ohno, 1981; Ohno and Epplen, 1993; Go, 1983). Such considerations make these Yockey-type calculations irrelevant!
The work done under the direction of Sauer has shown that the amino acid message "is highly degenerate in that many different sequences can code for proteins with essentially the same structure and activity." In other words "proteins are surprisingly tolerant of amino acid substitutions" (Bowie et al, 1990). "There is a wide range in tolerance to amino acid substitutions...The results reveal the high level of degeneracy in the information that specifies a particular protein fold." (Reidhaar-Olson and Sauer, 1990.) Bowie and Sauer (1989) describes the experimental technique used in these studies. This degeneracy would greatly facilitate the step-by-step evolution of proteins under the direction of natural selection. Thus we do not need to rely on an undirected search as Pandas assumes.
As the family of serum proteases illustrated, many proteins are derived from others by gene duplication, gene modification and exon shuffling (Blake, 1983; Doolittle, 1981; Furie and Furie, 1988; Neurath, 1984; Patthy, 1985). These mechanisms for evolving new genes and proteins from old are neglected by Pandas. Instead, they follow Ambrose and fall back upon supernatural events (see Pandas, Chapter 2). Totally different and unique proteins can arise through a frame-shift mutation. An actual example involves a new enzyme which arose around 1975 and allowed a microorganism Flavobacterium to break down the byproducts of nylon manufacture (Thwaites, 1985; Ohno, 1984; Okada et al, 1983). This event was made more likely by the fact that the original protein originated as a series of oligomeric repeats and retained a degree of internal repetitiousness (Ohno, 1984).
Again, Pandas' line of thinking (and its fallaciousness) can be illustrated by an analogy. Imagine that an exhaustive study restricted exclusively to the design, manufacture and operation of a modern jumbo jet concludes that such an airplane could only be made by a huge industrial complex. Furthermore, it is inconceivable that any airplane (past or present) could be simpler in design and still function, and certainly not so simple that it could be fabricated by two bicycle repairmen in the back of their shop! (If all the proteins must have originated in their modern forms, the original primordial cell must have had the complexity of a modern cell and there has been no evolution at the biochemical and intracellular level. Presumably Pandas believes there has been no ‘progressive creation' at these levels either.) The Wright brothers must have built a jumbo jet!
Conclusion
Pandas says, "Any view or theory of origins must be held in spite of unsolved problems; proponents of both design and unplanned descent acknowledge this." But there is a fundamental difference between these two views in regard to this point. Evolution is a theory of a specific natural mechanism. The theory not only recognizes its problems but specifies how they are to be solved, i.e. it serves as a guide to future research and has been extremely fruitful in serving this function. In contrast, the hypothesis of intelligent design simply postulates a totally mysterious, supernatural entity and provides no insight whatsoever as to how to solve its problems. (At various places in this book, Pandas says that intelligent design has its problems. But Pandas never says specifically what they are or how they are to be solved.)
References
Anon. 1980. The End of a Chemical Dichotomy. Mosaic 11(1): 23-28. (January/February.)
Awbrey, F. and W. Thwaites. 1981. Evolution vs. Creation. Aztec Lecture Notes. San Diego State University.
Blake, C. 1983. Exons—present from the beginning? Nature 306: 535-537.
Bowie, J. U., J. F. Reidhaar-Olson, W. A. Lim and R. T. Sauer. 1990. Deciphering the Message in Protein Sequences: Tolerance to Amino Acid Substitutions. Science 247: 1306-1310. (16 March.)
Bowie, J. U. and R. T. Sauer. 1989. Identifying determinants of folding and activity for a protein of unknown structure. Proceedings of the National Academy of Science USA. 86(7): 2152-2156. (April).
Doolittle, R. F. 1981. Similar Amino Acid Sequences: Chance of Common Ancestry? Science 214: 149-159.
Davie, E. W. and O. D. Ratnoff. 1964. Waterfall Sequence for Intrinsic Blood Clotting. Science 145: 1310-1312. (18 September).
Dayhoff, M. O., C. M. Park and P. J. McLaughlin. Building a Phylogenetic Tree: Cytochrome C. In: M. O. Dayhoff (Editor) 1972. Atlas of Protein Sequence and Structure 1972. Silver Spring, MD: National Biomedical Foundation. volume 5, pp.7-16.
Furie, B. and B. C. Furie. 1988. The Molecular Basis of Blood Coagulation. Cell, 53(4): 505-518 (May 20.)
Go, M. 1983. Modular structural units, exons, and function in chicken lysozyme. Proceedings of the National Academy of Sciences USA. 80(7): 1964-1968. (April.)
Jacob, F. 1977. Evolution and Tinkering. Science 196: 1161-1166. (10 June).
Lau, K. and K. A. Dill. 1989. A Lattice Statistical Mechanics Model of the Conformational and Sequence Spaces of Proteins. Macromolecules. 22: 3986-3997.
McFarlane, R. G. 1964. An Enzyme Cascade in the Blood Clotting Mechanism, and its Function as a Biochemical Amplifier. Nature 202: 498-499. (May 2).
Neurath, H. Evolution of Proteolytic Enzymes. Science 224: 350-357. (27 April).
Ohno, S. 1981. Original domain for the serum albumin family arose from repeated sequences. Proceedings of the National Academy of Sciences USA. 78(12): 7657-7661. (December.)
Ohno, S. 1984. Birth , of a unique enzyme from an altern, ative, reading frame of the preexisted internally repetitio, us coding sequence. Proceedings of the National Academy of Sciences USA. 81(8): 2421-2425. (April.)
Ohno, S. and J. T. Epplen. 1993. The primitive code and repeats of base oligomers as the primordial protein-encoding sequence. Proceedings of the National Academy of Sciences USA. n80(11): 3391-3395. (June.)
Okada, H. S. Negoro, H. Kimura and S. Nakamura. 1983. Evolutionary adaptation of plasmid-encoded enzymes for degrading nylon oligomers. Nature 306: 203-206.
Patthy, L. 1985. Evolution of the Proteases of Blood Coagulation and Fibrinolysis by Assembly from Modules. Cell 41: 657-663. (July).0
Reidhaar-Olson, J. F. and R. T. Sauer. 1990. Functionally Acceptable Substitutions in Two alpha-Helical Regions of Lambda Repressor. Proteins: Structure, Function, and Genetics. 7: 306-316.
Thwaites, W. M. 1985. New Proteins Without God's Help. Creation/Evolution 5(2): 1-3.
Yockey, H. P. 1977. A Calculation of the Probability of Spontaneous Biogenesis by Information Theory. Journal of Theoretical Biology. 67: 377-398.
(from Frank Sonleitner's critique of Of Pandas and People)